Digestion and Absorption
With your knowledge of secretions, anatomy, and directionality it is time to discuss digestion and absorption. As the food is passing through the gastrointestinal tract, with the help of muscle contractions and sphincters, it mixes with various secretions so that it can be digested. It then comes into contact with the epithelial tissue lining the gastrointestinal tract so that it can be absorbed into the body.
Learning Outcomes
In this section, you will learn…
- Digestion and absorption of macromolecules including carbohydrates, proteins, and lipids.
- Absorption of ions, minerals, vitamins, water, calcium and phosphate.
- The phases of gastric secretion.
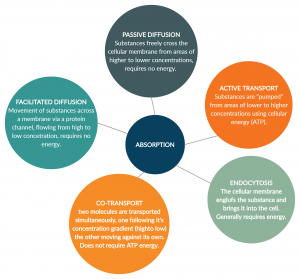
Take some time now to review the processes of absorption within the body talked about thus far as can be seen in figure 1. Keep these in mind when further reading about macromolecule digestion and absorption within this subchapter.
Carbohydrates
Various secretions in different organs, starting in the mouth, break down carbohydrates into their simplest forms prior to absorption. Carbohydrates play an extremely important role in the diet, and they are a direct source of glucose. Additionally, glucose is used as the primary fuel for our metabolism, which is why it is crucial that our body has adequate glucose levels. Being comfortable with the different mechanisms of both absorption and digestion and different classes of carbohydrates is important for understanding the mechanisms of GI function.
General Overview: Carbohydrate Digestion and Absorption
Before jumping into detail, take a look at this video to orient yourself to the bigger picture. This video is below course level, and can be used as a refresher, as well as to be used to think about additional detail you may have learned during class. This is a refresher of cell types from the salivary section within your notes, and additionally CCK from the small intestine portion of your notes. Integration can be explained and applied with knowledge learned in this video with focus on the time frame: 0:00-2:55.
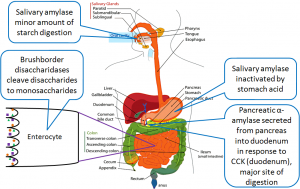
Take a look at figure 2 below which is a visual representation of the video and review what you just watched.
Enzyme category |
Enzyme name |
Source |
Substrate |
Product |
| Salivary enzyme | Salivary amylase | Salivary glands | Polysaccharides | Disaccharides and trisaccharides |
| Pancreatic enzyme | Pancreatic amylase | Pancreatic acinar cells | Polysaccharides | Disaccharides and trisaccharides |
| Brush border enzyme | Sucrase | Small intestine | Sucrose | Glucose and fructose |
| Brush border enzyme | Lactase | Small intestine | Lactose | Glucose and galactose |
| Brush border enzyme | Maltase | Small intestine | Maltose | Glucose |
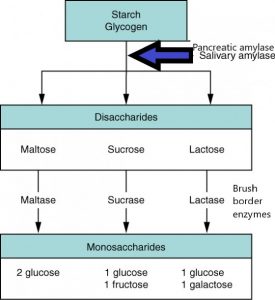
All these enzymes play a specific role in the process of carbohydrate digestion. See the following image to orient yourself to where the enzymes fit in:
Once digested down to their monomeric forms, carbohydrates must be absorbed by enterocytes. Table 2 complements the schematic in your notes and essentially describes how monosaccharides are taken up through the apical membrane of the enterocyte, via various transporters, then absorbed into the blood via the GLUT2 transporter, where they travel to the liver.
Food |
Breakdown products |
Absorption mechanism |
Entry to blood stream |
Destination |
| Carbohydrates | Glucose | Co-transport with sodium ions | Capillary blood in villi | Liver via hepatic portal vein |
| Carbohydrates | Galactose | Co-transport with sodium ions | Capillary blood in villi | Liver via hepatic portal vein |
| Carbohydrates | Fructose | Facilitated diffusion | Capillary blood in villi | Liver via hepatic portal vein |
As carbohydrates enter the mouth, digestion begins immediately with the release of salivary amylase. This enzyme is mixed with the bolus of food and travels down through the stomach into the small intestine. Once carbohydrates reach the small intestine, there is additional amylase secreted from the pancreas. The amylase secreted from both the mouth and the pancreas will break the alpha 1-4 bonds of polysaccharides (large carbohydrate molecules), breaking them down into disaccharides (small pieces of carbohydrates). Lactose, maltose, and sucrose are disaccharides found in the lumen of the small intestine, however, the epithelial cells cannot absorb them until they are broken down once more.
The small intestine, specifically the duodenum and jejunum, contain carbohydrate digesting enzymes along their brush border. Each disaccharide has a specific brush border enzyme that will convert them into their monomeric form. For example, lactose is broken down by lactase to make 1 glucose and 1 galactose. Maltose is broken down by maltase to make 2 glucose and sucrose is broken down by Sucrase to make 1 glucose and 1 fructose. They can then be absorbed by transporters into the epithelial cell. Galactose and glucose enter the cell via GLUT2, which is a symporter. GLUT2 also brings 2 sodium ions into the epithelial cell. Fructose is brought into the cell via GLUT5. This is a uniport, meaning GLUT5 does not bring in any other ions with the transport of fructose.
On the side of the epithelial cell, there is a sodium-potassium ATPase. It is important to not overlook it, as it actually has a very important role. It pumps sodium ions out of the epithelial cell, therefore lowering the concentration of sodium in the cell. This increases the drive to bring sodium into the epithelial cell, which can be accomplished by the GLUT2 symporter. As you already know, the GLUT2 symporter will then also bring glucose and galactose into the epithelial cell. Recall Na+/K+ ATPase and consider how the cell is set up and the relevance and sodium. How would a sodium gradient be maintained?
Try the following questions below to test your knowledge!
Protein
Protein digestion begins in the stomach. As you’ll learn, many of these proteins need to be activated prior to utilization. As a whole, protein digestion occurs in the stomach and the small intestine. Enzymes involved in protein digestion must be activated strategically, hence the very specific mechanism. This is important in the prevention of the digestion of tissues. A good summary of the process and enzymes involved in protein digestion is seen in Table 3. However, the details of these enzymes are beyond the course level. What is important is that you understand where they are all coming from, and what causes the activation of these enzymes.
Enzyme |
Inactive form |
Site of production |
Site of action |
Activated by |
Action |
End products |
| Pepsin | Pepsinogen | Chief cells | Stomach | Low pH | Cleaves proteins (polypeptides) | Peptides |
| Enteropeptidase | N/A | Cells of jejunum and duodenum | Small intestine | N/A | Activates trypsinogen | Trypsin |
| Trypsin | Trypsinogen | Pancreas | Small intestine | Enteropeptidase and self-activated | Activates proteases and cleaves proteins (polypeptides) | Active proteases, peptides |
| Proteases (various) | Zymogens | Pancreas | Small intestine | Trypsin | Cleaves proteins (polypeptides) | Peptides and amino acids |
| Peptidases (various) | N/A | Brush border | Small intestine | N/A | Cleaves peptides | Amino acids and oligopeptides |
Protein digestion begins in the stomach, where pepsinogen is secreted. Pepsinogen is quickly activated by the acidic environment of the stomach and becomes its active form, pepsin. Pepsin cleaves large proteins (also known as polypeptides), breaking them down into peptides. These peptides then travel to the small intestine to be further digested and absorbed.
There are numerous protein-digesting secretions accumulating in the small intestine. These secretions can be split into two production sites: the pancreas and the cells of the small intestine. As you know from the secretions section, the secretions from the pancreas are called zymogens, and they are all produced and secreted in an inactive form. A key zymogen that is secreted from the pancreas is trypsinogen. Trypsinogen is activated (to form trypsin) with the help of enteropeptidase. Since enteropeptidase is secreted from the cells of the small intestine, trypsinogen can only be activated once it exits the pancreatic duct and is in the duodenum. Once formed, trypsin can then go and activate all the other zymogens secreted from the pancreas. Once zymogens are activated, they are call proteases. The job of proteases is to break down polypeptides into peptides. It can be noted that some proteases break down polypeptides into amino acids, but most of the time another enzyme is needed.
Test Your Knowledge
Clinical Application:
You are a pediatrician and your patient’s parents tell you that despite eating full, nutrient-rich meals, their child is still hungry. The patient has Cystic Fibrosis, which can result in a buildup of mucus in the pancreatic duct. How would a buildup of mucus in the pancreatic duct have an effect on pancreatic secretions? Explain to the patient’s parents why their child is always hungry. Hint: How does the pancreatic duct help the function of digestion and absorption?
The final enzyme needed for digestion before absorption can take place is found in the brush border of the small intestine. Peptidases function to break down peptides into oligopeptides/dipeptides/tripeptides and amino acids. At this point, protein has been broken down enough that it can be absorbed.
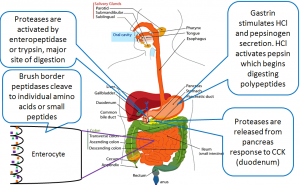
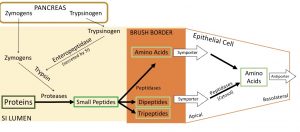
Take a look at figure 4 below which is a visual representation of protein digestion and absorption. Recall that enteropeptidase activates trypsinogen to produce trypsin. Then trypsin goes on to activate many other proteases.
Try the following question below to test your knowledge!
The absorption of proteins involves several different kinds of transporters. Amino acids can be brought into the cell via a uniport or a symporter. Dipeptides and tripeptides are brought into the cell via a symporter. On the apical side of the epithelial cell, there is also an antiporter maintaining the sodium and proton gradients for the aforementioned symporters. Also helping maintain the gradient is a sodium-potassium ATPase located on the side of the epithelial cell. Note that the antiporter and sodium-potassium ATPase are not included in the diagram below. Once in the epithelial cell, any dipeptides and tripeptides are broken down further to amino acids by a peptidase. The singular amino acids are transported over the basolateral side of the epithelial cell and into circulation via antiporter. This antiporter which amino acids out of the epithelial cell, and sodium in.
Try the following questions below to test your knowledge!
Lipids
Lipid digestion differs from that of carbohydrate and protein digestion, simply because lipids are hydrophobic. This section will summarize how lipids are digested and taken up by the body to either be used immediately or stored for later use.
Lipids have unique chemical properties that largely differ from carbohydrates and proteins. Fats are absorbed much differently, and throughout digestion lipid molecules are broken into smaller units by 3 main enzymes: lingual, gastric, and pancreatic lipase. The table below summarizes the function and properties of these three enzymes.
Enzyme category |
Enzyme name |
Source |
Substrate |
Product |
| Salivary enzyme | Lingual lipase | Lingual glands | Triglycerides | Free fatty acids, monoglycerides and diglycerides |
| Gastric enzyme | Gastric lipase | Chief cells | Triglycerides | Free fatty acids and monoglycerides |
| Pancreatic enzyme | Pancreatic lipase | Pancreatic acinar cells | Triglycerides that have been emulsified by bile salts | Free fatty acids and monoglycerides |
Lingual lipase and gastric lipase work in the stomach to start breaking down smaller lipids. However, most of the digestion of lipids occurs once they have reached the small intestine, where they are mixed with pancreatic lipase and bile. At the level of the intestine, these molecules must be kept apart so that they can be further digested and absorbed and this is the job of bile! The video below explains the general information about bile and bile salts, watch from 4:40-5:44.
The main job of bile in the small intestine is to emulsify lipids. Emulsification of lipids is the breakdown of large droplets of fat into smaller, easily digestible lipid droplets. This facilitates the job of lipases, as they can break the small lipid droplets into fatty acids. Free fatty acids can diffuse easily across the brush border into the epithelial cells. However, not all lipid droplets are broken down into fatty acids by lipases. Bile acids will also emulsify large lipid droplets, turning them into monoglycerides and long-chain fatty acids. This will then be packaged into a micelle. Micelles make contact with the brush border, and the lipids packaged in them diffuse out into the epithelial cell. The rest of the micelle, along with the bile salts used to break down and package the lipids, are left behind in the lumen of the GIT.
Once all the lipids have made their way into the epithelial cell, they need to be repackaged again. The long-chain fatty acids and monoglycerides that diffused out of the micelle combine to make triglycerides. These triglycerides, along with fatty acids, phospholipids, and cholesterol that have been absorbed, are packaged into chylomicrons. This repackaging happens in the Golgi network of the epithelial cell. An important feature of chylomicrons is that their outer surface is coated with proteins and hydrophilic phospholipid heads. Chylomicrons are essentially a package of lipids all wrapped up so they are non-lipid soluble, which facilitates transport throughout the body. After being repackaged, chylomicrons exit the epithelial cell on the basolateral side by exocytosis. The movement of lipids does not end here. Before we continue, review emulsification and the forms of lipid packaging by reading the following.
The visual representation of the emulsification and absorption of lipids, seen in figure 6, is at the level of the small intestine and enterocyte. Alongside is an image showing the structure of triglycerides and their digestion products, but note that the image does not mention chylomicron processing through the smooth ER. It is also specific to the absorption of only short-chain fatty acids and monoglycerides but neglects the alternative digestion of some long-chain fatty acids (Na+ symporter).
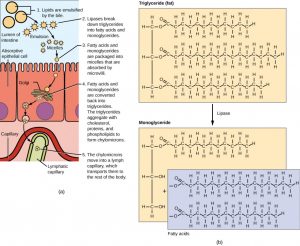
In all aqueous environments, lipids need to be “packaged” so that they don’t emulsify out of solution. As lipids are absorbed, moving from external to the internal environment, they change “packaging” forms too! Micelles package lipids in the external environment (the lumen), whereas chylomicrons package lipids in the cell. By forming an emulsion (in the chyme), bile salts increase the available surface area of the lipids many folds. The bile salts surround long-chain fatty acids and monoglycerides forming tiny spheres called micelles. The micelles move into the brush border of the small intestine absorptive cells where the long-chain fatty acids and monoglycerides diffuse out of the micelles into the absorptive cells leaving the micelles behind in the chyme. The long-chain fatty acids and monoglycerides recombine in the absorptive cells to form triglycerides, which aggregate into globules and become coated with proteins. These large spheres are called chylomicrons. [5]
One aspect of lipid absorption that makes it very different from carbohydrate and protein digestion is where it goes after being absorbed into the epithelial cell. As you previously read, lipids are packaged into chylomicrons and leave the epithelial cell via exocytosis. But where do they go from here?
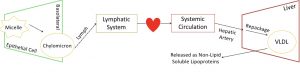
Proteins and carbohydrates are small and can be absorbed through epithelial cells straight into the bloodstream. However, chylomicrons are far too large to move directly into the blood, so they move into the lymph instead. Recall from earlier in this unit, structure of the GIT, that the lymph is found within the lamina propria of each villi. The lymph from the villi will meet up with the lymphatic system, just like how many capillaries come together to form a vein. The lymphatic system circulates around the body and will eventually meet up with the circulatory system. The contents of the lymph are then pumped out through systemic arteries. This means that while lipids aren’t directly absorbed into the blood, they do reach the circulatory system eventually.
The lymphatic system joins the circulatory system at the superior vena cava, a vein that empties directly into the heart. Once in the bloodstream, lipids are finally sent to the liver. Here they are re-packaged once again, but this time they form VLDL (very low-density lipoprotein), LDL (low-density lipoprotein) and HDL (high-density lipoprotein). These lipoproteins differ from lipids but are similar to chylomicrons because they are non-lipid soluble due to their protein coating.
Try the following question below to test your knowledge!
Ions
Ion absorption is reliant upon electrical gradients between cells. Take a look at figure 7below which illustrates how these electrical gradients work.
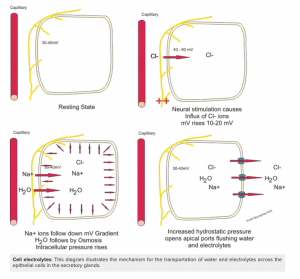
Before we look at the ion movement across epithelial cells, go back and look at sodium movement in diagrams depicting carbohydrate/glucose absorption and protein/amino acid absorption. You will notice that with the absorption of these nutrients, sodium is also being brought into the cell via a symporter. This starts a chain of events that ultimately results in the desired ion concentrations across the epithelial cell.
The first step is sodium moving into the cell during the absorption of glucose and amino acids. Once glucose is in the epithelial cell, it will be transported out the side of the same epithelial cell by a Na+/K+ ATPase. Remember way back to the start of the semester, Na+/K+ ATPase is a form of active transport, it requires ATP and can move ions against their concentration gradients. Now look at the diagrams of absorption of major nutrients again and notice the Na+/K+ ATPase can be seen on the side of the cell, and not on the apical or basolateral edges. This is important because we want to achieve a high sodium concentration in-between epithelial cells for the movement of other ions. So in summary, sodium enters the epithelial cell on the apical side with the absorption of nutrients like glucose and amino acids, then is transported out into space in between epithelial cells.
To examine chloride ion movement, let’s briefly switch gears from concentration gradients and now think about electrical gradients, another topic from the beginning of the course. Since the concentration of sodium in between cells is so high, it makes this area very positively charged. Since chloride ions are negatively charged, they will want to move down the electrical gradient towards sodium. This means chloride will move across the apical side of the epithelial cell directly into space in-between the cell.
Now let’s switch back to concentration gradients. We already know that water follows sodium, with the goal of keeping equal concentrations of each. Since the concentration of sodium is so high in-between cells, water will want to go there as well. Water moves through the apical side of the epithelial cells to the very same space in-between that has a high concentration of sodium. Let’s think about what the lumen of the GIT would look like now. Sodium, chloride, and water have all been absorbed into the body. This leaves potassium ions, which have become quite concentrated after all that water was absorbed. Once again, the ion will want to move from an area of high concentration to low, so naturally, potassium ions cross the apical side of the epithelial cell and are absorbed.
Try the following question below to test your knowledge!
Vitamins
Vitamins can either be water or fat-soluble — hydrophilic and hydrophobic molecules have different methods of transport into the body for use. The small intestine absorbs the vitamins that occur naturally in food and supplements. Fat-soluble vitamins (A, D, E, and K) are absorbed along with dietary lipids in micelles via simple diffusion. This is why you are advised to eat some fatty foods when you take fat-soluble vitamin supplements. Most water-soluble vitamins (including most B vitamins and vitamin C) also are absorbed by simple diffusion. An exception is vitamin B12, which is a very large molecule. Intrinsic factor secreted in the stomach binds to vitamin B12, preventing its digestion and creating a complex that binds to mucosal receptors in the terminal ileum, where it is taken up by endocytosis. [6]
Tips From Past Students
The paragraph above is beyond course level. What is important for course content is to be able to differentiate between fat and water solubles routes to absorption.
The absorption of fat-soluble vitamins and water-soluble vitamins are very different. Think back to the absorption of fats vs the absorption of water. They definitively don’t follow the same process, so neither does the absorption of fat-soluble vitamins and water-soluble vitamins. Water-soluble vitamins are absorbed simply by diffusion or mediated transport. They can pass into the epithelial cells quite easily. Fat-soluble vitamins on the other hand do not simply diffuse into the epithelial cell. They are packaged into micelles, just like lipids are. From here, their absorption follows the same route as the absorption of lipids.
Water
Each day, about nine liters of fluid enter the small intestine. About 2.3 liters are ingested in foods and beverages, and the rest is from GI secretions. About 90% of this water is absorbed in the small intestine. Water absorption is driven by the concentration gradient of the water: The concentration of water is higher in chyme than it is in epithelial cells. Thus, water moves down its concentration gradient from the chyme into cells. As noted earlier, much of the remaining water is then absorbed in the colon. Remember that water is following sodium through the GIT. [7]
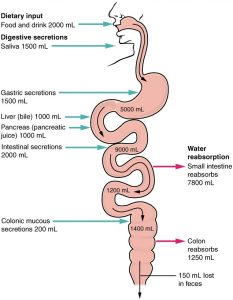
It is recommended that people drink 2L of water a day to be hydrated, but in reality, the GIT needs between 8 and 9L of water for it to function! So where does the rest of it come from? The majority of this section has been centered around the absorption of various nutrients, but to understand the absorption of water we must also discuss its secretion. The GIT needs roughly 8.5L of water. If we say 2L of that was ingested from you drinking water, that still leaves 6.5L. This extra 6.5L of water actually is secreted from various organs throughout the GIT to help it function. The GIT secretes 6.5 liters of water per day, which supplements that 2L that are ingested. Specifically, saliva secretes 1.5 L, gastric secretes 2.0 L, the pancreas secretes 2.0 L, the liver secretes 0.5 L, and the small intestine secretes 1.0 L. However, the large intestine does not contribute to the secretion of water because, at that point in the GIT, the only goal is to complete the absorption of water.
Now that we have the necessary amount of water for the GIT to function, it can be absorbed. Water is absorbed in two locations: the small intestine and the large intestine. The majority of the water (approximately 6.5L) is absorbed in the small intestine. Remember we discussed water absorption briefly in the ion absorption section. Water is absorbed in the small intestine as it moves along its concentration gradient (i.e. it has a much lower concentration in the epithelial cells than in the lumen). It also moves in the same direction as sodium ions, which are frequently absorbed in conjunction with other nutrients. In the large intestine, approximately 1.9L of water is absorbed, and the remaining 100mL is excreted.
Tips From Past Students
Water absorption is not so much a component of the GIT unit, rather it is focused more on throughout the kidney unit. This bit of information is just to compliment the notes that we have on H2O flux near the end of the unit
Calcium
Calcium is an important micronutrient used to maintain bone health and basic cellular processes. This section will summarize how the body maintains calcium homeostasis. Calcium is a key electrolyte as 99% is deposited in the bones and the remainder is associated with hormone release and cell signaling. The plasma concentration of Ca2+ is 2.2 mmol/l, and phosphate is 1.0 mmol/l.
Vitamins D’s role in calcium metabolism includes being responsible for controlling the absorption of calcium and increased amounts of vitamin D result in increased amounts of calcium absorption. The mechanism behind this is by increasing components involved in calcium absorption such as calcium channels, calbindin, and sodium-calcium pumps. It helps to control the balance between plasma and bone.
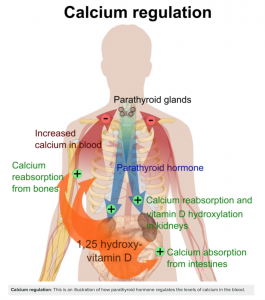
Parathyroid hormones also have a role in calcium excretion. What stimulates parathyroid hormone includes monitoring blood calcium which can indicate if levels are too low, and if calcium levels get low parathyroid hormone levels to increase. This helps to control the balance between plasma and bone. The goal of parathyroid is to increase blood calcium. The mechanisms behind this include increased resorption of calcium from bone, decreased excretion of calcium and upregulation of vitamin D, which will ultimately increase calcium reabsorption in the small intestine [8]
You have learned about calcium in past units, however, the focus here is on the mechanism of its absorption. We will look at how the body takes in calcium from the lumen of the GIT into the epithelial cell. Recall from the hormones unit that when calcium levels are low in the plasma, this causes an increase in PTH and an increase in levels of 1,25-(OH)2-D. Thus far, all you know is that this results in an increase in calcium absorption in the small intestine (as well as other processes in the kidney and bone, but that is not the focus here).
1,25-(OH)2-D enters the epithelial cell by simple diffusion. It acts on the nucleus to increase the synthesis of various proteins that are needed to absorb calcium. On the apical (luminal) side, calcium channels are needed for calcium to be able to enter the cell. The work of 1,25-(OH)2-D doesn’t end here, as calcium still needs to be transported across the basolateral side of the cell and into the blood. Once in the epithelial cell, calcium must be bound to a protein called calbindin. As you already know, calcium is often used as a second messenger in various signaling pathways. Leaving calcium unbound in an epithelial could result in the initiation of unwanted processes. So for the cell’s protection, calcium binds to calbindin. To get calcium across the basolateral side of the cell, two proteins are used. The first is an exchanger, which transports calcium out of the cell while transporting 2 sodium ions into the cell. The second is a calcium pump that, with the use of ATP, transports calcium out of the cell while simultaneously transporting a proton into the cell. It is important to note that calcium is released from calbindin once it is ready to exit the epithelial cell.
Try the following question below to test your knowledge!
In summary, 1,25-(OH)2-D increases the synthesis of key proteins that help calcium go from the lumen of the GIT, across the apical side into the epithelial cell, then across the basolateral side into the blood. These proteins are calcium channels, calbindin, calcium exchanger, and calcium pump. Consider how calcium absorption would impact whole-body calcium homeostasis, refer to figure 10 for some guidance.
Phosphate
The absorption of phosphate is similar to the absorption of calcium, where 1,25-(OH)2-D also affects the amount of phosphate that is absorbed. Once again, 1,25-(OH)2-D can diffuse into the epithelial cell and increase the synthesis of proteins that help get phosphate from the lumen of the GIT into the body. Phosphate crosses the apical side of the epithelial cell with the help of a symporter, which also brings in 2 sodium ions. To maintain the gradient needed for this symporter, a sodium-potassium ATPase is located on the side of the epithelial cell. The movement of phosphate across the cell is not well known yet and is beyond the level of this course. What is important is to know that there is some kind of transporter that brings phosphate across the basolateral side of the epithelial cell and into the body. In summary, 1,25-(OH)2-D acts on the nucleus of epithelial cells to increase the synthesis of proteins needed for phosphate absorption, just as it did for calcium. These proteins are the phosphate symporter, sodium-potassium ATPase, and the phosphate transporter (basolateral side). Try the following question below to test your knowledge!
Test Your Knowledge
Clinical Application:
You’re a doctor and one of your patients comes in with excruciating stomach pains. After a few tests, you diagnose her with a stomach ulcer which is caused by the propagation of helicobacter pylori in the stomach. This bacteria eats away at the stomach lining. Using your knowledge of the GIT, explain why this bacteria’s presence in the stomach is problematic and why this ulcer has formed. What foods do you recommend that your patient stay away from if they are being treated for a stomach ulcer? You administer antibiotics to kill H. pylori as well as drugs to reduce the amount of acid released into the stomach. Which pumps are being blocked with the use of the drug? While this is necessary for the healing of the stomach lining, what are the consequences of these gastric acid-lowering drugs on digestion and absorption? Hint: Think of the function of the stomach lining and what problems it would cause if it weren’t there.
Phases of Gastric Secretions
There are various phases throughout the GIT that are triggered by the presence of food in various areas of the body. As you will learn, the stimulation of each of these three phases triggers the release of different secretions in different areas of the body that promote the digestion of foodstuffs. The secretions in the GIT occur in three phases: the cephalic phase, the gastric phase, and the intestinal phase. Watch this whole video to orient yourself with these phases.
“Phases of Acid Gastric Secretion- Physiology Medical Animation” by Dr.G. Bhadu Prakash is All Rights Reserved.
The phases are stimulated by different factors, however, they all result in stimulation of gastric secretions as demonstrated in the figure below.
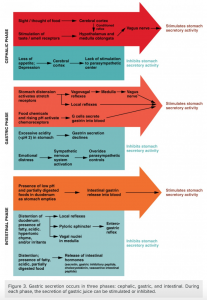
The cephalic phase (reflex phase) of gastric secretion, which is relatively brief, takes place before food enters the stomach. The smell, taste, sight, or thought of food triggers this phase. For example, when you bring a piece of sushi to your lips, impulses from receptors in your taste buds or the nose are relayed to your brain, which returns signals that increase gastric secretion to prepare your stomach for digestion. This enhanced secretion is a conditioned reflex, meaning it occurs only if you like or want a particular food. Depression and loss of appetite can suppress the cephalic reflex.
The gastric phase of secretion lasts 3 to 4 hours and is set in motion by local neural and hormonal mechanisms triggered by the entry of food into the stomach. For example, when your sushi reaches the stomach, it creates distention that activates the stretch receptors. This stimulates parasympathetic neurons to release acetylcholine, which then provokes the increased secretion of gastric juice. Partially digested proteins, caffeine, and rising pH stimulate the release of gastrin from enteroendocrine G cells, which in turn induces parietal cells to increase their production of HCl, which is needed to create an acidic environment for the conversion of pepsinogen to pepsin, and protein digestion. Additionally, the release of gastrin activates vigorous smooth muscle contractions. However, it should be noted that the stomach does have a natural means of avoiding excessive acid secretion and potential heartburn. Whenever pH levels drop too low, cells in the stomach react by suspending HCl secretion and increasing mucous secretions.
The intestinal phase of gastric secretion has both excitatory and inhibitory elements. The duodenum has a major role in regulating the stomach and its emptying. When partially digested food fills the duodenum, intestinal mucosal cells release a hormone called intestinal (enteric) gastrin, which further excites gastric juice secretion. This stimulatory activity is brief, however, because when the intestine distends with chyme, the enterogastric reflex inhibits secretion. One of the effects of this reflex is to close the pyloric sphincter, which blocks additional chyme from entering the duodenum. [9]
Key Takeaways
Consider the following concepts to help guide your studies:
- How digestion and absorption of each macromolecule take place.
- How absorption of ions, minerals, vitamins, water, calcium, and phosphate occurs.
- Understand the role of each phase of gastric secretion.
Subchapter Quiz
The questions below can be used to assess your knowledge within this chapter. There are multiple-choice questions that you should attempt without referring to your notes. The questions will provide you with responses to your answers to guide your studying but should not be used as your only resource.
Media Attributions
- ABSORPTION-1 © OpenStax adapted by Ingrid Jones is licensed under a CC BY (Attribution) license
- Review of Carbohydrate Digestion © Brian Lindshield is licensed under a CC BY (Attribution) license
- carbsteps © Donna Browne adapted by Ingrid Jones is licensed under a CC BY (Attribution) license
- Review of Protein Digestion © Brian Lindshield is licensed under a CC BY (Attribution) license
- Protein Digestion Absorption Flowchart © Alexa King is licensed under a CC BY (Attribution) license
- Lipid and Triglyceride Digestion © Charles Molnar and Jane Gair is licensed under a CC BY (Attribution) license
- Lipid Absorption © Alexa King is licensed under a CC BY (Attribution) license
- Electrolyte Transport © Boumphreyfr is licensed under a CC BY-SA (Attribution ShareAlike) license
- Digestive Secretions and Absorption of Water © Lumen Learning is licensed under a CC BY (Attribution) license
- Calcium Regulation © Mikael Haggstrom is licensed under a Public Domain license
- Phases of Secretion © Lumen Learning is licensed under a CC BY (Attribution) license
- 23.7 chemical digestion AND Absorption: A closer look - anatomy and physiology. (n.d.). Retrieved April 19, 2021, from https://openstax.org/books/anatomy-and-physiology/pages/23-7-chemical-digestion-and-absorption-a-closer-look ↵
- 23.7 chemical digestion AND Absorption: A closer look - anatomy and physiology. (n.d.). Retrieved April 19, 2021, from https://openstax.org/books/anatomy-and-physiology/pages/23-7-chemical-digestion-and-absorption-a-closer-look ↵
- Rye, C., Avissar, Y., Jurukovski, V., Fowler, S., Wise, R., Roush, R., . . . Gair, J. (2015, May 14). 15.3 digestive system processes. Retrieved April 19, 2021, from https://opentextbc.ca/biology/chapter/15-3-digestive-system-processes/ ↵
- 23.7 chemical digestion AND Absorption: A closer look - anatomy and physiology. (n.d.). Retrieved April 19, 2021, from https://openstax.org/books/anatomy-and-physiology/pages/23-7-chemical-digestion-and-absorption-a-closer-look ↵
- Rye, C., Avissar, Y., Jurukovski, V., Fowler, S., Wise, R., Roush, R., . . . Gair, J. (2015, May 14). 15.3 digestive system processes. Retrieved April 18, 2021, from https://opentextbc.ca/biology/chapter/15-3-digestive-system-processes/ ↵
- 23.7 chemical digestion AND Absorption: A closer look - anatomy and physiology. (n.d.). Retrieved April 18, 2021, from https://openstax.org/books/anatomy-and-physiology/pages/23-7-chemical-digestion-and-absorption-a-closer-look ↵
- 23.7 chemical digestion AND Absorption: A closer look - anatomy and physiology. (n.d.). Retrieved April 18, 2021, from https://openstax.org/books/anatomy-and-physiology/pages/23-7-chemical-digestion-and-absorption-a-closer-look ↵
- Boundless. (n.d.). Boundless anatomy and physiology. Retrieved April 18, 2021, from https://courses.lumenlearning.com/boundless-ap/chapter/electrolyte-balance/ ↵
- Learning, L. (n.d.). Anatomy and physiology. Retrieved April 18, 2021, from https://courses.lumenlearning.com/nemcc-ap/chapter/the-stomach-and-the-pancreas/ ↵
An enzyme in saliva that separates carbohydrates into smaller molecules.
An enzyme produced by the pancreas that is released into the small intestine to breakdown carbohydrates.
An enzyme that promotes the breakdown of sucrose to glucose and fructose.
An enzyme that promotes the breakdown of lactose to galactose and glucose.
An enzyme that facilitates the breakdown of maltose to two glucose molecules.
Cells that line small and large intestine surfaces, otherwise known as the intestinal absorptive cells.
Enlargement due to an increase in internal pressure.
Hormone-producing cells located in the GIT, pancreas, and stomach.

{kind=link}
{kind=link}